Herbertia 53: 129-144 (1998)
PROGENY OF HIPPEASTRUM PAPILIO
John D. Fellers
2290 Longwood Drive,
Auburn AL 36830-7126, U.S.A.
資料來源:International Bulb Society
譯者:小亨利 (Henry) 2011.01.25
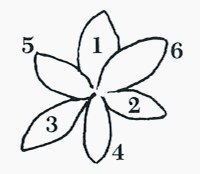
花瓣部分的編號為 1-2-3,依順時針旋轉,1號是最上層的花瓣部分。
花萼部分的編號為 4-5-6,以順時針旋轉; 4號是最下層的花萼部分。
鳳蝶和巴西其他一些原生孤挺,最重要一項特徵是兩片編號5和6的花萼特別寬而明顯,編號4的主花萼卻是最狹窄的。一般來說,2號和3號花瓣都比4號花萼稍寬 。這些花瓣通常是狹長的橢圓形。1號花瓣的寬度較編號5和6的花萼,約窄 10mm。這與荷蘭育種的經典型leopoldii孤挺,是很明顯不同的新特徵。荷蘭育種的孤挺,一般1號花瓣最寬 4號花萼最窄,但其他花瓣寬度都較平均,不像鳳蝶孤挺花的差異那麼大。 因此這成為評量鳳蝶子代育種時的重要指標。
鳳蝶孤挺花的兩片編號5和6的花萼特別寬而花紋顏色對比明顯 像極了蝴蝶,因此被稱為鳳蝶 (papilio 拉丁語的意義就是蝴蝶)。
《請參考圖一》她那暗紫的色脈絡和斑點在1號花瓣及5、6的花萼上特別加深,襯上淡綠到黃綠的底色,搶眼的色彩對比效果絕佳,誕生了令人 "耳目一新" 的花朵。
一般孤挺及其他石蒜花的共同特徵是2號和3號花瓣,無論花紋顏色和大小,通常是相似而左右對稱的。5和6號的花萼也是相似而左右對稱。1號花瓣的花紋色彩會比較接近5和6號的花萼。 4號花萼花紋色彩則比較接近2號和3號花瓣的肋下方,然而2號和3號肋上方的花色與脈絡就比較接近5和6號的花萼。
鳳蝶不例外也遵循這種模式,在花的上半部顯現出強烈突出而沉重的暗栗色脈紋。 4號花萼則幾乎全為淺綠色(有些株會呈現黃綠色)有一定比例的後代會出現一絲絲淡紅色脈絡,4花萼肋的兩側。 2號和3號花辦肋下方會比較接近4號花萼的色彩與花紋,但2號和3號花辦肋上方的花色與紋路會比較接近5和6號的花萼。
剛入門的鳳蝶孤挺花育種者首要工作,就是認識這些鳳蝶孤挺花的特殊基本特質。以下將會陸續討論到:
|
1、美國現有的鳳蝶似乎都是自交不親合,就連對一些兄弟株也是如此。但是對血緣較遠的鳳蝶就沒有這問題。鳳蝶似乎是對交配的對象很挑,必須是與它相容的類型。這裡實驗所
用的鳳蝶也是如此。 無論是與老品種或是與荷蘭園藝種雜交,最初授粉後似乎有了結果,但發展28日後就突然中止《譯者按:這可能是由於二倍體與四倍體雜交的原因(鳳蝶是二 倍體,荷蘭園藝種是四倍體。)》。 其中一些種子會繼續成長進入成熟的階段,但最後都是發育不良的空殼。運氣好偶爾可以得到3-4顆可播種的種子,令人不禁十分期待他們的長大與開花。相對的一些少數的鳳蝶F1和F2後代,就顯得不會那麼的挑剔講究。 結論是鳳蝶很難配種,除了靠毅力和適當的條件外,往往還需要靠運氣才能找到合適的配種。 2、鳳蝶的開花期很固定,很少有太大變異。在我種了25年鳳孤的經驗裡,很少遇到過了正常花期還會開花的情形。然而 F1 and F2子代的變異性就較大了,鳳蝶在此地溫室的花期約是三月第一周的前後十天,依當季天氣狀況而稍有變動,花期可延續約三周(在加州此花經常於春秋兩季時,年開兩次)。 為了達到使鳳蝶有足夠種類的混種後代,我們必須建立一個花粉銀行。大部分和鳳蝶可以交配的品種,花季都是在秋冬季尾隨鳳蝶的開花期。因此當以鳳蝶為母本時,父本就必須靠上季保留下來的花粉來進行雜交。所幸孤挺的花粉很耐儲藏,在室溫乾燥的環境下保存期長達數週。旺季的新鮮花粉都裝在有標示的花粉罐,存放於冰箱之中。旺季之後這些花粉罐就會被蓋起來並放入雙層袋中,在放置於冰箱中冷藏(千萬別放於冷凍庫)。孤挺花粉雖然耐儲藏,但也必須在舊花粉失效前,經常更新新鮮花粉。 3、鳳蝶基本上是終年常綠,只於仲夏約有一段為期3~4週的休眠期後,就會開始另一季新的成長。此時是移植換盆的好時機,但是鳳蝶一旦根系被干擾後,必須等到根部重新生長後才會開花。因此換盆通常會導致花期延後。和其他孤挺一樣,鳳蝶根系爆盆後比較容易來花。 4、 純種鳳蝶,似乎對一些孤挺病毒有免疫力。此性狀也會遺傳至一些後代但不是全部,例如鳳蝶(母本)與原生孤挺 puniceum(父本) 的後代,對於父本容易得的一些病毒,也是沒有免疫力。 但此類後代往往很早就會開花,播種三年後就會開花。不必等到一般的4~7年才會來花,甚至有二年實生苗就開花,或一年F1苗當年就來花,但很可能因為選錯父本而未結子。 5、我們在此地,沒有做過任何鳳蝶或其後代的耐寒性測試。這些孤挺都是養殖於溫室中,直到春季霜害解除之後,才會移出溫室。十一月初霜後立即移入溫室保護,有一次30°F(-1°C )的寒流突然來襲,有一批盆栽還來不及放入溫室,只受到一些簡單的覆蓋保護。沒想到溫度居然突然降到24°F(-4°C )並維持了數小時,純種鳳蝶完全不受影響,在下一季也正常開花。其他雜交種也沒受到太大影響,但是有一顆種在四加侖桶裡的 H. papilio x H. fragrantissimum 卻受到重創 (在同樣的保護狀態下) 之後,就趕緊放入溫室保護。有些球失去所有葉子,下一季無法正常開花,這情形延續了數年。 6、強健的生命力是鳳蝶的一項寶貴特質,其球根相當巨大,此特質是顯性會遺傳給後代。鳳蝶的花朵遺傳特質則是隱性,與其他種的F1雜交後代中,顯示花色完全受其他親本影響 。 花的形狀及大小或許還可以見到鳳蝶的遺傳,但花色幾乎可以肯定完全受到其他親本的控制。 若是不知道其他親本的背景,很難在其F1雜交後代中,目視出鳳蝶的特徵。F2雜交後代也是如此,除非是回交鳳蝶厚才會有例外 (這並不容易 比預期的還難達成)。F3後代中,雙親都有鳳蝶血統的子代才會出現一些鳳蝶特徵,(有極少數則是很接近的)。 當鳳蝶是母本時父本的花色會重現,有部分會出現類似鳳蝶的花型,但顏色品質及斑紋都改變了。看起來就不像真的鳳蝶,但有少數會比較接近鳳蝶,底色可能是較淡或較深的綠色。棕紅色的斑紋和斑塊也會有變化,顏色會偏向褐色而不是強勢的棕紅色。 花型並沒有像典型鳳蝶一般,兩片編號5和6的花萼特別寬而明顯。若和鳳蝶並列,則可以明顯看出5和6號花萼的差異。 茲將一些鳳蝶後代 以鳳蝶為母本回交的F1及包括其衍生(F2,F3)的雜交後代,整理描述如下: 以鳳蝶為親本導入的園藝品種,一般被用來測量的典型部位有:花朵直徑 (即從1號花瓣尖到4號花萼尖端 約為19cm),1號花瓣寬是4.4cm,2和3號花瓣寬是3.3cm,l和4號寬是3.4cm 最顯著的5和6花萼寬則是5.8cm。此株鳳蝶親本是褐栗色(非猩紅色)斑塊及脈絡疊在淺綠的底色上。花藥裂開後是綠色,厚實的花梗約68公分高。 第一次的突破是發現與鳳蝶可以雜交的另一巴西原生種 H. aulicum (1977)。 此原生孤 植株球根都比鳳蝶小一號,花直徑卻是16.5cm,這顆 H. aulicum 看起來像1966由Robert Goedert引進的 H. aulica Ker Gawl幾乎是黑色的斑紋,夾在藍綠色的喉部與深猩紅的花瓣之中。 花藥如鳳蝶一樣是綠色,兩片編號5和6的花萼,特別寬而明顯,2、3和4號花瓣比較狹長。 [下列雜交種皆依照傳統方式標示親本,先標母本,再標父本。] <請參考圖七> 鳳蝶 母本 X H. aulicum 父本 在此簡稱 Papaul (就是我們俗稱的紅鳳蝶),這項雜交於1977完成,於1978年又再進行一次。1977完成的幼苗被標示為Papaul 77-1,Papaul 77-2,77-3 等等,這些子代的球根都較其雙親大。雖然色調有些深淺差異,但多數的Papaul 77 大致都是深紅底色。 就像是大朵的 H.aulicum (除了一個顏色較淺以外),有些如石榴紅,有些則是暗紅色。 <請參考圖十一> Papaul 78的花就更深紅了,有些花是暗紅色含深紫色條紋,花瓣比 Papaul 78的普遍狹長。 所有的子代都擁有顯著寬廣的5和6花萼,這是其雙親共有的特質。 紅鳳蝶的雙親開花期不同,鳳蝶是三月一號,H. aulicum 傳統上都是十一月來花。紅鳳蝶花直徑約19cm,壯碩的花梗約63cm高,深綠茂盛的葉子具有抗病毒特性。綠色花藥的特性被遺傳下來,紅鳳蝶的後代是傳承鳳蝶特徵的重要品系。 下一個雜交種,就是與上述親本相反的,H. aulicum x 鳳蝶父本,簡稱 Aulpap 其他巴西原生種也可以批配 Hippeastrum corriensis 和 H. ackermanii,都可以和鳳蝶雜交。 但其中 Aulpap 77-1 發展出像,香殊蘭似的巨大球根及充 滿強健生命力的葉子。 他的開花性,穩定而可靠。 每季開出接近 ± 20cm 的花朵 ,母球與子球生出3~4梗花 。如其雙親一樣擁有顯著寬廣的5和6花萼,其他花辦則較為細瘦。 看起來和一些Papaul 78很像, Aulpaps 的花色比 Papauls 淡許多,偏向暗橘紅色(非猩紅色)。 他的花瓣脈絡分明,與一些 Papaul 78 很像。 其後代包括兄弟株,截至目前為止,只能被 H. angustifolium 的花粉交配。 其他的雜交包括自交都是失敗的。 鳳蝶和在佛羅里達舊農場買的 H. puniceum 很容易雜交,花色是杏橘色黃色花喉短肋植株和花都比典型佛州生長的 H. puniceum 來的大顆。雜交種 80-2 (H. papilio x H. puniceum-belladonna type)的子代,花色由淡橘色到杏桃色,黃喉黃肋,和父本有很大不同。 其中一顆是可愛的整片亮橘色,花比其他兄弟株小與其父本大小相同。其他杏桃色黃喉的子代的花辦,都比父本寬大而重疊。這些子代特別是那顆可愛的亮橘花,都被用來做為育種的材料。 H. papilio 和 H. corriensis 雜交的後代,讓人很難預測其結果。 孤挺corriensis 屬迷孤,球根相當小,約蒜頭那麼大 11.4公分。花是橘紅色帶綠喉,中間是亮綠色,花瓣瘦長,通常六月底開花。其雜交的後代通常比他原種的開花期還早。H. papilio 和 H. corriensis 無論誰當母本或父本,雜交的後代植株都界 於親本大小之間,花型較小但較瘦長,編號5和6的花萼沒有特別的顯著。 Papcor 81-1 (H. papilio x H. corriensis) 的花呈現瓷白底色,暗紅色(到紫紅色)的纖細或寬脈條紋,喉部及下肋都是深綠色。親本相反的 Corpap 81-3 (H. corriensis x H. papilio) 則花多暗紅較少白色,花的下半部出現深綠色,下肋明顯的紫紅色充滿花的中央及編號5和6的花萼,一直延伸至喉部。 鳳蝶與 ackermannii雜交後代 H. papilio and H. x ackermannii (H. aulicum x H. x johnsonii) 呈現與 鳳蝶和 H. corriensis雜交後代類似的結果;這些後代都不像其親本 然而以鳳蝶為母本的栽培種,花色比以鳳蝶為父本 (H. x ackermannii為母本)的顏色來的更淡 孤挺 ackermannii 與其母本 aulicum 差不多大小 花瓣比較張不開 只有12.5cm寬,花的顏色是似紅葡萄酒般的酒橙色。 <請參考圖四> 鳳蝶與 ackermannii雜交後代 簡稱 Papacker (H. papilio x H.ackermanii) 81-5 的花色近乎整片的深紅色,只有些許的白線脈紋。 2、3號花瓣的肋與尖端顏色較淡,色彩帶點紫褐色,相鄰的是羽化的綠色肋,其喉部幾乎是黑色的。這是平均標準,有些當然更亮或更暗,其5和6號的花萼都特別寬而顯著。 <請參考圖十> 親本相反的 Ackerpap 81-6 (H. x ackermanii x H. papilio) 產生更豐富的暗紅寶石花色。其中被選定作為展示例子的植株,顯現了淺色的花邊,只有少量的後代花瓣比 Papacker 81-5 更寬。 以 Ackerpap81-6作母本與其他非鳳蝶的種間雜交,都會產生醒目的深寶石紅色的花朵。 孤挺 Ackerpap 無論採父本或母本方式雜交,都已被證明是一個重要的親本。 <請參考圖二> 有一株以 Ackerpap為母本 以 'Sumac Pinini' 為父本的後代及其兄弟株,產生顏色獨特而接近父本的栽培種,我們把它標示為 85-Apap-18 (Ackerpap x 'Sumac Pinini')此株的兄弟株展現了不同深淺的花色。 其中最深色的 85-Apap-18a <請參考圖十六> 花直徑是16cm 花是褐栗色帶白邊,葉子都比鳳蝶更藍一點。大多數的 85-Apap-18的花色比較淡,且有細條紋或斑點的深色豹斑,此項特徵溯源其父本。子代的花大小各異由14.4公分至19.5公分不等。這些更小更淡的花就被標記為 85-Apap-18b。他們的花朵大小和形式與 85-APAP-18a 並無顯著不同。花朵較大時,花瓣就相對也越細長橢圓。 <請參考圖九> 鳳蝶與豹紋雜交的後代,似乎花色都是斑點而且顏色加深 89-Pap-101 (H. neopardinum x H. papilio) 產生的後代與85-Apap-18b。顏色較淡的後代開花非常類似,但他的花朵有非常清晰的細紋,那些花瓣細長的栽培種 被命名為'Spotty'。 鳳蝶,據聞是目前市面上至少一種以上,黃花孤挺的遺傳親本。 其對黃花親本的遺傳影響力,可藉由鳳蝶與黃花的F1子代 85-Pap-27 (H. papilio x H. mananita)中,展露無遺。黃孤挺 mananita Doran 是中型花,花型和目前市面流行的建國小黃 Germa 很相像。 他的花型是較長的喇叭狀,並且花色更趨黃鶯色,葉姿與生長方式都大不同。 <請參考圖十五> 85-Pap 27的 F1子代由父本 Mananita 遺傳了鶯黃底色,由母本鳳蝶遺傳了鳳蝶其他子代都有的共同特徵 美麗的斑紋 對親本的了解有助於此項雜交的成功。鳳蝶與 Hippeastrum papilio x (Korsakoff form of H. greenii x H.aglaiae) 雜交後代產生了顯性的白底 在5和6號花瓣上出現紅色條紋 細細的紅邊與綠喉 形成強烈的對比有趣的是,這個栽培種和 H. papilio x H. fragrantissimum 的顏色神似。 但是,第一花瓣與第四花萼都拉得特別長,所以整體花朵看起來是不尋常的大。 花的寬度也很有趣,5和6的花萼特別的顯著令人想起鳳蝶的身影。 此株被命名為 84-Pap-4 [H. papilio x (Korsakoff form of H. greenii x H. aglaiae)]。 |
 |
| 圖一、Hippeastrum papilio #23 | |
 |
|
| 圖二、Ackerpap x 'Sumac' | |
 |
|
| 圖三、H. papilio x H. aulicum | |
 |
|
| 圖四、H. papilio x H. x ackermannii | |
 |
|
| 圖五、H. papilio x H. fragrantissimum | |
 |
|
| 圖六、H. papilio x H. striatum | |
 |
|
| 圖七、H. papilio x H. aulicum | |
 |
|
| 圖八、(H. papilio x H. fragrantissimum) x Sprekelia formosissima | |
 |
|
| 圖九、H. neopardinum x H. papilio | |
 |
|
| 圖十、Ackerpap 81-6 | |
 |
|
| 圖十一、Papaul 78-1 x Papfrag | |
 |
|
| 圖十二、H. iguazuannum x Austpapst | |
 |
|
| 圖十三、Hippeastralia 'Mystique' x Sprekelia | |
 |
|
| 圖十四、Papfrag x double | |
 |
|
| 圖十五、Papaul 78-1 x (Papaul 77-3 x H. mananita) | |
 |
|
| 圖十六、85-Apap-18a |
<請參考圖六> 鳳蝶的紅斑部分其實帶有棕色,而鳳蝶 Hippeastrum papilio x H. striatum 雜交後代,帶出了棕色。 這似乎是這種親本組合的趨勢,這裡採用的父本 H. striatum是杏橘色細辦花,85-Pap-7 (H. papilio x H. striatum) 後代產生的花色是由棕橘色至喉部有些微條紋的亮橘色。 另一個後代 85-Pap-8,有類似的親本,是2,3號花瓣及4號花萼有條紋的中型花,沒有特別顯著的花瓣或花萼。喇叭長筒狀有時長度比花朵還大,記錄的比例約是花的筒徑 19cm 比花的直徑 17cm花會微微下垂,這是 H. striatum的特質。 花梗初期是紫紅色隨著開花期間轉為綠色。
92-Pap-11 是試驗'Mystique'雜交鳳蝶子代的結果,其後代充滿強健生命力,這是典型的鳳蝶後裔特色。 花張的很開約16.7cm 花瓣寬度都一致。 1號花瓣約比5、6號花萼寬 0.6cm,4號花萼比其他四個花瓣約窄 1.2cm。顏色是鮮亮橘紅色重疊於純黃底之上。 是一款很 "陽光型" 的孤挺。 此栽培種的花色很特別,亮紅色斑紋取代了原本鳳蝶的咖啡紅斑紋部分,而且斑紋密度顯得更紮實。 原來黃綠色的底色也變成純黃色,進一步與第二代和第三代鳳蝶的 交叉雜交下 開創了更廣闊鳳蝶世界的序幕。強健生命力似乎在這群活潑子代中不斷提升 因而產生了更寬長的葉子,更寬厚的花梗( 有時是更高)就連大花也很常見。
我們觀察到,更多變化的兄弟苗株,由單一花房中誕生了。原本與鳳蝶雜交不親和的品種,也顯示能和某些鳳蝶的F1和F2子代授粉結子。 例如: Papauls(鳳蝶 X aulicum),鳳蝶 80-2 ( 鳳蝶 X puniceum )。
鳳蝶與勁香孤挺的雜交後代 H. PAPILIO x H. FRAGRANTISSIMUM
我購買了一顆鳳蝶與勁香孤挺的雜交後代,來加入我的育種計劃。為了協助維持親本及栽培種的紀錄,我們簡稱其為 Papfrag,與其他的鳳蝶交配種一樣。鳳蝶與勁香孤挺的後代 有白底色與紅線條及白肋交錯一起喉部和肋部,都還展示著代表貴族的淡黃綠色 但是喇叭筒的長度比花朵本身還長 約17.2cm 花梗長度約有90cm以上, 此栽培種的側球所開的花 會與母球的特殊斑紋產生密度與厚度上的變異,這是非常有趣的現象,同時來自父本遺傳的影響 會帶有一點宜人的香味。
84-Pap-20 (Papaul x Papfrag) 是由兩個與鳳蝶雜交過的F1子代,再雜交的後代。兄弟株開的花,花色變異度很大,由幾乎全白到深紅(近黑色)都有。其中有象牙白綠喉的花,顯示父本的影響,花色刷過一點燻衣草的藍色調。白花後代的 84-Pap-20a 顯示了鳳蝶的活力,花型及大小。5和6號花萼寬而顯著,只有這兩片花萼才有,些微紅色纖細的斑紋覆蓋於白底上(必須非常接近觀察);同時這兩片花萼外圍鑲有細紅邊。其他花瓣都是純白,花朵直徑約18cm。
84-Pap-20b 的花是屬暗紅色,比較接近父本 Papfrag。 花瓣寬度平均。 花只比H. gracilis 稍大朵 花徑 14。8cm。在持續延伸鳳蝶與aulicum 的F1雜交種(Papauls)的育種過程中,我們發現了許多珍貴如寶石的後代 90-Pap-25 [Papaul 78-2 x (Papaul 77-3 x H. mananita)] 此花具有鳳蝶的強健生命力,花直徑約18.4cm 5,6號的花萼也是寬而明顯,花色是獨特的甜瓜綠色刷上淡鮭魚的色斑 90-Pap-28,有著相同的親本背景,花長的幾乎是一樣,只是色斑是刷上淡粉紅色,花色是衡量 Papaul 77-3 x H. mananita。淡色花改良育種的主要標準,其暗色系的後代擁有,獨特濃郁的深紅木色(給人一種高貴的氣質)。 這高雅的顏色在與其他顏
色匹配中,經常脫穎而出。純橘的 80-2 (H. papilio x H. puniceum-belladonna type) 也產生了不少人讚嘆的後代,8427-Auspapst擁有 8。25 吋 可愛黃喉橘色花;他是由 Papilio 80-2與澳洲進口的孤挺雜交而成的。 8427-Auspapst 至目前為止還未結子過,但其花粉還是有生殖能力的。
<請參考圖十二> Hippeastrum iguazuanum x 8427-Auspapst 雜交產生了 90-AM-1開出了 25。2cm的大花 厚實的花梗支持著沉重的花朵 1號花瓣約有 8.3cm 寬;其他兩片花萼約 7、6cm寬。 花有遺傳到一定程度的垂花型,花張的很開,完全看不到 H. iguazuanum 花瓣反捲成鉤狀的特徵,明亮強烈的橘色被淡黃肋貫穿直至花瓣尖端。
<請參考圖五> 三種與 H. papilio x H. fragrantissimum (Papfrag) 的 F1 雜交種展現出此栽培種的育種多樣性。由園藝栽培發展出來的鑲邊品種,被用來做為母本與 Papfrag 父本雜交後代。 94-PAPP-21 (鑲邊種) x Papfrag 顯示出純鑲邊, 沒有任何斑點或污點,高雅淡綠的底色取代了原有的瓷白色。他也同時遺傳了鳳蝶的強健生命力 85cm 高的花梗直徑 3.8cm。 寬而顯著的花萼 讓19.2cm的花朵 整體顯得寬扁 花瓣寬而重疊。 1號花瓣約 9.6cm 寬。 2,3號花瓣約 9.3cm 寬; 4號花萼約 5.8cm 寬 其他兩片花萼約 7.4cm寬。 有點直立的花萼 使花型看起來更有趣味。另一個花型較正常的兄弟株,產生了一個完全潔白如瓷的花朵,襯上深色寶石紅的鑲邊。
<請參考圖十四> 93-Pap-13 (Papfrag x 覆瓣孤挺)產出大量的花瓣。共約有24瓣,93-Pap-13 也許是四倍體。花型明顯比父本大約21cm寬,花萼長約15cm 在花梗上添加不尋常的重荷。當兩朵花同時開放時,即使是高66cm的厚實花梗也需要額外的支撐。 此花的四片底辦上有著與 Papfrag (H. papilio x H. fragrantissimum)花萼上相同的淡象牙綠。上方的花瓣反映出 Papfrag 花萼的紋路,Papfrag的細鑲邊特質也呈現在此栽培種的全部花瓣上。
<請參考圖八> 屬間交配的 94-Papsk-14 (H. Papfrag x Sprekelia TMH) 提供了一個有趣的情景,這是 H. papilio x H. fragrantissimum 的雜交種為母本與燕子水仙為父本的跨屬雜交。後代都是展現強健生命力的植株,盡管植株及葉子與傳統孤挺的沒兩樣,但質地較厚,可以抵擋住強風吹襲。此花給人的深刻印象是長的喇叭型花萼及微開的花瓣,花朵含苞待放時,1號花瓣會輕微向上反捲,其他花瓣則仍然保持平直。這種特質不禁令人聯想到在其他跨屬(孤挺交燕子水仙)的雜交後代,含苞待放時都可以觀察到這項共同特點。雜交後代的花朵外型會還原成原來一般孤挺的模樣,花徑約 13.4cm 喇叭狀的花萼長達 11.5 cm。有寬而顯著的花萼。花色亮麗,清秀的洋紅底色配上一直延伸至花瓣中央的潔白花肋。
總結來說,這些都是鳳蝶雜交育種的優良案例。 同時也顯示鳳蝶的後代能為我們石蒜科的育種舞台上,增加了不少的經典劇目。 然而育種的路途還很遙遠,很多拓荒者還在前線繼續為大家努力奮鬥。
REFERENCES
- Baker, J.G. 1888. Description of the Amaryllideae. Handbook of the Amaryllideae: subgenus Omphalissa (H. aulicum Herb., App 31. A. aulica Gawl.); repr. 1934. Year Book Amaryllis Society, 1: 38, 42.
- Blossfeld, Harry. 1970. Two Brazilian Amaryllis Species-Amaryllis Aulica Ker. Gawler. Plant Life 26: 58-62.
- Blossfeld, Harry. 1973. Amaryllis Calyptrata Ker-Gawl. Plant Life 29: 30-34.
- Buchmann, Fred J. 1969. Hybridizing with Amaryllis Species -1960. Plant Life 25: 79-82.
- Caldwall, Sam. 1968. Amaryllis aulica Ker Gawl. Plant Life 24: 36-39.
- Cardenas, Martin. 1960. New Bolivian Amaryllis (A. fragrantissima). Plant Life 16: 32-33.
- Cardenas, M. 1972. New Bolivian Species (A. neoleopoldii Card). Plant Life 28: 52-54.
- Castillo, Jose A. 1986. Notes on the Cultivation of South American Bulbous Plants. Plant Life 42: 63-66, 68.
- Cothran, Charles Dewitt 1979. Yellow Flowered and other Amaryllis Hybrids. Plant Life 35: 61-65.
- Cothran, C.D. 1980. The Quest for Large Yellow Flowered Hybrids. Plant Life 36: 19-22.
- Cothran, C.D. 1980. A Fragrant Tri-Specific Amaryllis Hybrid. Plant Life 36: 81-82.
- Cothran, C.D. 1982. The Continuing Pursuit of Yellows. Plant Life 33: 61.
- Cothran, C.D., 1984. Large Yellow Amaryllis Hybrids. HERBERTIA 40: 105-111.
- Craft, Douglas D. 1962. Amaryllis x Johsonnii. Plant Life 18: 28-29.
- Deme, John Wade. 1979. Double Amaryllis Up-Date. Plant Life, 35: 66.
- Doran, J. L. 1972. Some Cultural Requirements of Amaryllis Species. Plant Life 28: 89-91.
- Goedert, Robert D. 1965. Amaryllid Culture-Amaryllis Species. Plant Life 21: 127-128.
- Hayward, Wyndham. 1934. The Mead Strain of Nehrling Hybrid Amaryllis. Year Book American Amaryllis Society 1: 62.
- Hayward, Wyndham. 1948. Hybrid Amaryllis Improvement. HERBERTIA 15: 63-68.
- Hayward, Wyndham. 1950. Amaryllis of Tomorrow. Plant Life 6: 97-104.
- Hayward, Wyndham. 1959. Old Amaryllis Clones. HERBERTIA 15: 31-35.
- Henry, Mary G. 1951. x Amaryllis Gladwynensis. Plant Life 7: 119.
- Howard, Thad M. 1990. The Genus Sprekelia in Mexico, Central America and South America. HERBERTIA 49: 97-100.
- Lobner, Max. 1937. Amaryllis in Germany. HERBERTIA 4: 186-187.
- Mann, Kenneth and Ed Pencall. 1980. Southern California Hemerocallis and Amaryllis Show. Plant Life 36: 35.
- Manning, Russell H. 1974. Sprekelia-Amaryllis Cross. Plant Life 30: 85-86.
- Manning, R.H. 1982. North Midland Amaryllid-Report 1982. Plant Life 38: 97-103,
- Meerow, Alan W., Timothy K. Brocht and Michael E. Kane. 1991. Hippeastrum Breeding at the University of Florida. HERBERTIA 47: 4-10.
- Mertzweiller, James K. 1968. Genetics and Breeding Amaryllis pseudopardina. Plant Life 24: 73-74.
- Mertzweiller, James K. 1969. An Amaryllis Breeding Project-1968 Report. Plant Life 25: 88-91.
- Pradham, Udai Chandra. 1970. An Apparent Sprekelia-Amaryllis Cross. Plant Life 26: 117.
- Ravenna, Pedro Felix. 1970. Contributions to South American Amaryllidaceae III—Amaryllis Papilio Rav. sp. nov.) Plant Life, 26: 82-84.
- Ravenna, P.F. 1971. Contribution to South American Amaryllidaceae IV—Amaryllis iguazuanna, Rav. sp. nov. Plant Life 27: 63-65.
- Rosoff, I.K., and W. Quinn Buck. 1970. Southern California Official Show. Plant Life 26: 19-20.
- Traub, Hamilton P. 1934. The Nehrling Hybrid Amaryllis. Year Book American Amaryllis Society 1: 61.
- Traub, H.P., 1946. Terminology for the Floral Envelope of Amaryllidaceae. HERBERTIA 13: 98.
- Wheeler, Willis H. 1968. Amaryllis striata for Rapid Increases. Plant Life 24: 111-1,12.
- Whitaker, Thomas W. 1982. Amaryllis Aulica Naturalized in Southern California. Plant Life 82: 38, 95-96.
- Williams, Paul H. Jr. 1970. An Aquatic Amaryllis Species from Argentina. Plant Life 26: 65.
- Wilson, Marcia Client. 1981. Amaryllid Hybrids of J.L. Doran. Plant Life 37: 109.